Признаки характеризующие митоз. Деление клеток. Митоз и мейоз, фазы деления. Этапы деления клетки
Фаза G1характеризуется возобновлением интенсивных процессов биосинтеза, который в период митоза резко замедляется, а на короткое время цитокинеза – прекращается вовсе. Общее содержание белка за время этой фазы увеличивается непрерывно. Для большинства клеток существует критическая точка в фазе G1 так называемая точка рестрикции. При ее прохождении в клетке происходят внутренние изменения, после которых клетка должна пройти все последующие фазы клеточного цикла. Границу между фазами S и G2 определяет появление вещества - активатора S-фазы.
Фаза G2 рассматривается как период подготовки клетки к началу митоза. Ее длительность меньше остальных периодов. В ней происходит синтез белков деления (тубулин) и наблюдается фосфорилирование белков, участвующих в конденсации хроматина.
Профаза
Во время профазы происходят два параллельных процесса. Это постепенная конденсация хроматина, появление отчетливо видимых хромосом и дезинтеграция ядрышка, а также формирование веретена деления, обеспечивающего правильное распределение хромосом между дочерними клетками. Эти два процесса пространственно разделены ядерной оболочкой, которая сохраняется в течение всей профазы и разрушается только в ее конце. Центром организации микротрубочек у большинства животных и некоторых растительных клеток служит клеточный центр или центросома. В интерфазной клетке он располагается сбоку от ядра. В центральной части центросомы располагаются две центриоли, погруженные в ее материал под прямым углом друг к другу. От периферической части центросомы отходят многочисленные трубочки, образованные белком тубулином. Они существуют и в интерфазной клетке, образуя в ней цитоскелет. Микротрубочки пребывают в состоянии очень быстрой сборки и разборки. Они нестабильны и их массив постоянно обновляется. Например, в клетках фибробластов в культуре in vitro среднее время жизни микротрубочек составляет менеее 10 мин. В начале митоза микротрубочки цитоплазмы распадаются, а затем начинается их восстановление. Сначала они появляются в околоядерной зоне, формируя лучистую структуру – звезду. Центром ее образования является центросома. Микротрубочки являются полярными структурами так как молекулы тубулина, из которых они образуются ориентированы определенным образом. Один конец ее удлиняется втрое быстрее других. Быстро растущие концы названы плюс концами, медленно растущие минус-концами. Плюс концы ориентированы вперед по направлению роста. Центриоль – это небольшая цилиндрическая органелла толщиной около 0,2 мкм и длиной 0,4 мкм. Ее стенку образуют девять групп триплетов трубочек. В триплете одна трубочка полная и две примыкающие к ней неполные. Каждый триплет наклонен в сторону центральной оси. Соседние триплеты соединены между собой поперечными сшивками. Новые центриоли возникают только путем удвоения уже существующих. Этот процесс совпадает со временем синтеза ДНК в S-фазе. В G1 периоде происходит раздвигание центриолей, образующих пару, на несколько микрон. Затем на каждой из центриолей, в ее средней части, под прямым углом строится дочерняя центриоль. Рост дочерних центриолей завершается в G2 фазе, но они еще погружены в единую массу центросомного материала. В начале профазы каждая пара центриолей становится частью отдельной центросомы, от которой отходит радиальный пучок микротрубочек – звезда. Сформировавшиеся звезды отодвигаются друг от друга по двум сторонам ядра, становясь впоследствии полюсами веретена деления.
Метафаза

Прометафаза начинается с быстрого распада ядерной оболочки на мембранные фрагменты, не отличимые от фрагментов ЭПС. Они сдвигаются к периферии клетки хромосомами и веретеном деления. На центромерах хромосом образуется белковый комплекс, который на электронных фотографиях выглядит как пластинчатая трехслойная структура – кинетохор. Обе хроматиды несут по одному кинетохору, именно к нему прикрепляются белковые микротрубочки веретена деления. Методами молекулярной генетики выяснено, что информация определяющая специфическую конструкцию кинетохоров заключена в нуклеотидной последовательности ДНК в районе центромеры. Микротрубочки веретена, прикрепленные к кинетохорам хромосом играют очень важную роль, они во-первых, ориентируют каждую хромосому относительно веретена деления так, чтобы два ее кинетохора были обращены к противоположным полюсам клетки. Во-вторых, микротрубочки перемещают хромосомы, чтобы их центромеры оказались в плоскости экватора клетки. Этот процесс в клетках млекопитающих занимает от 10 до 20 мин и завершается к концу прометафазы. Число микротрубочек, связанных с каждым кинетохором, различно у разных видов. У человека их бывает от 20 до 40, у дрожжей – 1. С хромосомами связываются плюс концы микротрубочек. Кроме кинетохорных микротрубочек веретено деления содержит еще полюсные микротрубочки, которые отходят от противоположных полюсов и на экваторе сшиваются специальными белками. Микротрубочки, которые отходят от центросомы и не включаются в веретено деления, называют астральными они образуют звезду.
Метафаза. Занимает значительную часть митоза. Она легко распознается по двум признакам: двухполюсной структуре веретена деления и метафазной хромосомной пластинке. Это относительно стабильное состояние клетки, многие клетки можно оставить в метафазе на несколько часов или дней, если их обработать веществами деполимеризующими трубочки веретена. После удаления агента митотическое веретено способно к восстановлению и клетка способна завершить митоз.
Анафаза

Анафаза начинается быстрым синхронным расщеплением всех хромосом на сестринские хроматиды, каждая из которых имеет свой кинетохор. Расщепление хромосом на хроматиды связано с репликацией ДНК в районе центромеры. Репликация такого небольшого участка происходит за несколько секунд. Сигнал к началу анафазы исходит из цитозоля, он связан с кратковременным быстрым повышением концентрации ионов кальция в 10 раз. Электронная микроскопия показала, что у полюсов веретена происходит скопление мембранных пузырьков, богатых кальцием. В ответ на анафазный сигнал сестринские хроматиды начинают движение к полюсам. Это связано сначала с укорочением кинетохорных трубочек (анафаза А), а затем – раздвигание самих полюсов,связанное с удлинением полярных микротрубочек (анафаза В). Процессы относительно самостотельны, на что указывает их разная чувствительность к ядам. У разных организмов вклад анафазы А и анафазы В в окончательное расхождение хромосом различен. Например, в клетках млекопитающих анафаза В начинается вслед за анафазой А и заканчивается, когда веретено достигает длины в 1,5-2 раза больше, чем в метафазе. У простейших анафаза В преобладает, в силу чего веретено удлиняется в 15 раз. Укорочение кинетохорных трубочек идет путем их деполимеризации. Субъединицы теряются с плюс конца, т.е. со стороны кинетохора, в результате кинетохор передвигается вместе с хромосомой к полюсу. Что касается полюсных микротрубочек. То в анафазе происходит их сборка и удлинение по мере расхождение полюсов. К концу анафазы хромосомы полностью разделяются на две идентичные группы на полюсах клетки.
Деление ядра и цитоплазмы связаны. Важную роль при этом играет митотическое веретено. В животных клетках уже в анафазе в плоскости экватора веретена появляется борозда деления. Она закладывается под прямым углом к длинной оси митотического веретена. Образование борозды обусловлено активностью сократимого кольца, которое располагается под мембраной клетки. Оно состоит из тончайших нитей – актиновых филаментов. Сократимое кольцо обладает силой, достаточной для того, чтобы согнуть тонкую стеклянную иглу, введенную в клетку. По мере углубления борозды толщина сократимого кольца не увеличивается, так как часть филаментов теряется при уменьшении его радиуса. После завершения цитокинеза сократимое кольцо полностью распадается, плазматическая мембрана в области борозды деления стягивается. Некоторое время в зоне контакта вновь образованных клеток сохраняется тельце из остатков тесно упакованных микротрубочек. В растительных клетках, имеющих жесткую клеточную оболочку, цитоплазма разделяется путем образования новой стенки на границе между дочерними клетками. В растительных клетках нет сократимого кольца. В плоскости экватора клетки формируется фрагмопласт, постепенно расширяющийся от центра клетки к ее периферии, пока растущая клеточная пластинка не достгнет плазматической мембраны материнской клетки. Мембраны сливаются, полностью разделяя образовавшиеся клетки.
Размножение клеток – один из важнейших биологических процессов, является необходимым условием существования всего живого. Репродукция осуществляется путем деления исходной клетки.
Клетка – это наименьшая морфологическая единица строения любого живого организма, способная к самопроизводству и саморегуляции. Время ее существования от деления до гибели или же последующей репродукции называется клеточным циклом.
Ткани и органы состоят из различных клеток, которые имеют свой период существования. Каждая из них растет и развивается, чтобы обеспечивать жизнедеятельность организма. Длительность митотического периода различна: клетки крови и кожи входят в процесс деления каждые 24 часа, а нейроны способны к репродукции только у новорожденных, а затем вовсе утрачивают способность к размножению.
Существует 2 вида деления — прямое и непрямое . Соматические клетки размножаются непрямым путем, гаметам или половым клеткам присущ мейоз (прямое деление).
Митоз — непрямое деление
Митотический цикл
Митотический цикл включает 2 последовательных этапа: интерфазу и митотическое деление.
Интерфаза (стадия покоя) – подготовка клетки к дальнейшему разделению, где совершается дублирование исходного материала, с последующим его равномерным распределением между новообразованными клетками. Она включает 3 периода:
- Пресинтетический (G-1) G – от английского gar, то есть промежуток, идет подготовка к последующему синтезу ДНК, выработка ферментов. Экспериментально проводилось ингибирование первого периода, вследствие чего клетка не вступала в следующую фазу.
- Синтетический (S) — основа клеточного цикла. Происходит репликация хромосом и центриолей клеточного центра. Только после этого клетка может перейти к митозу.
- Постсинтетический (G-2) или премитотический период — происходит накопление иРНК, которая нужна для наступления собственно митотического этапа. В G-2 периоде синтезируются белки (тубулины) – основная составляющая митотического веретена.
После окончания премитотического периода начинается митотическое деление . Процесс включает 4 фазы:
- Профаза
– в этот период разрушается ядрышко, растворяется мембрана ядра (нуклеолема), центриоли располагаются на противоположных полюсах, формируя аппарат для деления. Имеет две подфазы:
- ранняя — видны нитеобразные тела (хромосомы), они еще не четко отделены друг от друга;
- поздняя — прослеживаются отдельные части хромосом.
- Метафаза – начинается с момента разрушения нуклеолемы, когда хромосомы хаотично лежат в цитоплазме и только начинают двигаться к экваториальной плоскости. Между собой все пары хроматид связаны в месте центромеры.
- Анафаза – в один момент разобщаются все хромосомы и движутся к противоположным точкам клетки. Это короткая и очень важная фаза, поскольку именно в ней происходит точный раздел генетического материала.
- Телофаза – хромосомы останавливаются, снова образуется ядерная мембрана, ядрышка. Посередине образуется перетяжка, она делит тело материнской клетки на две дочерние, завершая митотический процесс. В новообразованных клетках снова начинается G-2 период.
Мейоз — прямое деление
 Мейоз — прямое деление
Мейоз — прямое деление
Существует особый процесс репродукции, встречающийся только в половых клетках (гаметах) – это мейоз (прямое деление) . Отличительной чертой для него является отсутствие интерфазы. Мейоз из одной исходной клетки дает четыре, с гаплоидным набором хромосом. Весь процесс прямого деления включает два последовательных этапа, которые состоят из профазы, метафазы, анафазы и телофазы.
Перед началом профазы у половых клетках происходит удвоение исходного материала, таким образом, она становится тетраплоидной.
Профаза 1:
- Лептотена — хромосомы просматриваются в виде тоненьких ниток, происходит их укорочение.
- Зиготена — стадия конъюгации гомологичных хромосом, как следствие образуются биваленты. Конъюгация важный момент мейоза, хромосомы максимально сближаются друг с другом, чтобы осуществить кроссинговер.
- Пахитена — происходит утолщение хромосом, их все большее укорочение, идет кроссинговер (обмен генетической информацией между гомологичными хромосомами, это основа эволюции и наследственной изменчивости).
- Диплотена – стадия удвоенных нитей, хромосомы каждого бивалента расходятся, сохраняя связь только в области перекреста (хиазмы).
- Диакинез — ДНК начинает конденсироваться, хромосомы становятся совсем короткими и расходятся.
Профаза заканчивается разрушением нуклеолемы и формированием веретена деления.
Метафаза 1 : биваленты расположены посередине клетки.
Анафаза 1 :к противоположным полюсам движутся удвоенные хромосомы.
Телофаза 1 :завершается процесс деления, клетки получают по 23 бивалента.
Без последующего удвоения материала клетка вступает во второй этап деления.
Профаза 2 : снова повторяются все процессы, которые были в профазе 1,а именно конденсация хромосом, что хаотично располагаются между органеллами.
Метафаза 2 : две хроматиды, соединенные в месте перекреста (униваленты), располагаются в экваториальной плоскости, создавая пластинку, названную метафазной.
Анафаза 2: — унивалент разделяется на отдельные хроматиды или монады, и они направляются к разным полюсам клетки.
Телофаза 2
: процесс деления завершается, формируется ядерная оболочка, и каждая клетка получает по 23 хроматиды.
Мейоз – важный механизм в жизни всех организмов. В результате такого деления мы получаем 4 гаплоидные клетки, которые имеют половину нужного набора хроматид. Во время оплодотворения две гаметы образуют полноценную диплоидную клетку, сохраняя присущей ей кариотип.
Сложно представить наше существования без мейотического деления, иначе все организмы с каждым последующим поколение получали бы удвоенные наборы хромосом.
Деление клетки - биологический процесс, лежащий в основе размножения и индивидуального развития всех живых организмов.
Наиболее широко распространенная форма воспроизведения клеток у живых организмов - непрямое деление, или митоз (от греч. «митос» - нить). Митоз состоит из четырех последовательных фаз. Благодаря митозу обеспечивается равномерное распределение генетической информации родительской клетки между дочерними клетками.
Период жизни клетки между двумя митозами называют интерфазой. Она в десятки раз продолжительнее митоза. В ней совершается ряд очень важных процессов, предшествующих делению клетки: синтезируются молекулы АТФ и белков, удваивается каждая хромосома, образуя две сестринские хроматиды, скрепленные общей центромерой, увеличивается число основных органоидов клетки.
Митоз
В процессе митоза различают четыре фазы: профазу, метафазу, анафазу и телофазу.
- I. Профаза - самая продолжительная фаза митоза. В ней спирализируются и вследствие этого утолщаются хромосомы, состоящие из двух сестринских хроматид, удерживаемых вместе центромерой. К концу профазы ядерная мембрана и ядрышки исчезают и хромосомы рассредоточиваются по всей клетке. В цитоплазме к концу профазы центриоли отходят к полосам и образуют веретено деления.
- II. Метафаза - хромосомы продолжают спирализацию, их центромеры располагаются по экватору (в этой фазе они наиболее видны). К ним прикрепляются нити веретена деления.
- III. Анафаза - делятся центромеры, сестринские хроматиды отделяются друг от друга и за счет сокращения нитей веретена отходят к противоположным полюсам клетки.
- IV. Телофаза - делится цитоплазма, хромосомы раскручиваются, вновь образуются ядрышки и ядерные мембраны. После этого образуется перетяжка в экваториальной зоне клетки, разделяющая две сестринские клетки.
Так из одной исходной клетки (материнской) образуются две новые - дочерние, имеющие хромосомный набор, который по количеству и качеству, по содержанию наследственной информации, морфологическим, анатомическим и физиологическим особенностям полностью идентичен родительским.
Рост, индивидуальное развитие, постоянное обновление тканей многоклеточных организмов определяется процессами митотического деления клеток.
Все изменения, происходящие в процессе митоза, контролируются системой нейрорегуляции, т. е. нервной системой, гормонами надпочечников, гипофиза, щитовидной железы и др.

Мейоз (от греч. «мейоз». - уменьшение) - это деление в зоне созревания половых клеток, сопровождающееся уменьшением числа хромосом вдвое. Он состоит и двух последовательно идущих делений, имеющих те же фазы, что и митоз. Однако продолжительность отдельных фаз и происходящие в них процессы значительно отличаются от процессов, происходящих в митозе.
Эти отличия в основном состоят в следующем. В мейозе профаза I более продолжительна. В ней происходит конъюгация (соединение) хромосом и обмен генетической информацией. (На рисунек вверху профаза отмечена цифрами 1, 2, 3, конъюгация показана под цифрой 3). В метафазе происходят те же изменения, что и в метафазе митоза, но при гаплоидном наборе хромосом (4). В анафазе I центромеры, скрепляющие хроматиды, не делятся, а к полюсам отходит одна из гомологичных хромосом (5). В телофазе II образуются четыре клетки с гаплоидным набором хромосом (6).
Интерфаза перед вторым делением у мейоза очень короткая, в ней ДНК не синтезируется. Клетки (гаметы), образующиеся в результате двух мейотических делений, содержат гаплоидный (одинарный) набор хромосом.
Полный набор хромосом - диплоидный 2n - восстанавливается в организме при оплодотворении яйцеклетки, при половом размножении.
Половое размножение характеризуется обменом генетической информации между женскими и мужскими особями. Оно связано с образованием и слиянием особых гаплоидных половых клеток - гамет, образующихся в результате мейоза. Оплодотворение представляет собой процесс слияния яйцеклетки и сперматозоида (женской и мужской гамет), при котором восстанавливается диплоидный набор хромосом. Оплодотворенную яйцеклетку называют зиготой.
В процессе оплодотворения можно наблюдать различные варианты соединения гамет. Например, при слиянии обеих гамет, имеющих одинаковые аллели одного или нескольких генов, образуется гомозигота, в потомстве которой сохраняются все признаки в чистом виде. Если же в гаметах гены представлены различными аллелями - образуется гетерозигота. В ее потомстве обнаруживаются наследственные зачатки, соответствующие различным генам. У человека гомозиготность бывает лишь частичной, по отдельным генам.
Основные закономерности передачи наследственных свойств от родителей к потомкам были установлены Г. Менделем во второй половине XIX в. С этого времени в генетике (науке о закономерностях наследственности и изменчивости организмов) прочно утвердились такие понятия, как доминантные и рецессивные признаки, генотип и фенотип и др. Доминантные признаки - преобладающие, рецессивные - уступающие, или исчезающие в последующих поколениях. В генетике эти признаки обозначаются буквами латинского алфавита: доминантные обозначаются заглавными буквами, рецессивные — строчными. В случае гомозиготности каждая из пары генов (аллелей) отражает либо доминантные, либо рецессивные признаки, которые в обоих случаях проявляют свое действие.
У гетерозиготных организмов доминантная аллель находится в одной хромосоме, а рецессивная, подавляемая доминантом, в соответствующем участке другой гомологичной хромосомы. При оплодотворении образуется новая комбинация диплоидного набора. Следовательно, образование нового организма начинается со слияния двух половых клеток (гамет), образующихся в результате мейоза. Во время мейоза происходит перераспределение генетического материала (рекомбинация генов) у потомков или обмен аллелями и их соединение в новых вариациях, что и определяет появление нового индивида.
Вскоре после оплодотворения происходит синтез ДНК, хромосомы удваиваются, и наступает первое деление ядра зиготы, которое осуществляется путем митоза и представляет собой начало развития нового организма.
Профаза
. В профазе происходит конденсация хромосом, и они становятся видимыми при световой микроскопии. Хромосомы по мере компактизации ДНП приобретают строение хорошо окрашивающихся нитей. Число хромосом равно 4n, что соответствует количеству ДНК 4с. В связи с инактивацией генов в области ядрышкового организатора и угнетением синтеза РНК в профазе отмечается исчезновение ядрышек. Ядерная оболочка постепенно распадается на фрагменты и мелкие мембранные пузырьки. При этом к противоположным полюсам клетки расходятся центриоли.
В сателлитном участке материнской центриоли
начинается образование микротрубочек, из которых формируются нити веретена деления.
Метафаза
. Характерным событием для метафазы является перемещение хромосом в экваториальную плоскость веретена. Здесь они располагаются строго закономерно, образуя метафазную пластинку (при взгляде на веретено деления сбоку). Если рассматривать группу метафазных хромосом со стороны полюсов веретена, то отчетливо выступает фигура, напоминающая звезду (так называемая материнская звезда). В этот период можно определить число, форму и размеры хромосом (d-хромосом, двойных хромосом), составляющих метафазную пластинку.
К концу метафазы
продольные половинки хромосом (сестринские хроматиды) обособляются на всем протяжении, кроме зоны первичной перетяжки.
Для каждого вида животных характерно строго постоянное число хромосом в соматических клетках. Для человека оно равно 46. По длине хромосом различают чередование окрашенных и неокрашенных участков. При этом каждая хромосома отличается неповторимым рисунком дифференциальной окраски. Хромосомы человека подразделяются на 7 групп по их размерам и особенностям строения (А, В, С, D, Е, F, G) и каждая хромосома имеет свой номер. Совокупность признаков строения хромосом, их размеров и числа составляет то, что называют кариотипом.
Анафаза включает процесс расхождения хромосом к полюсам делящейся клетки. Механизм движения хромосом объясняется гипотезой скользящих нитей, согласно которой состоящие из микротрубочек нити веретена, взаимодействуя друг с другом и с сократительными белками, тянут хромосомы к полюсам. Скорость движения хромосом достигает 0,2-0,5 мкм/мин, а вся анафаза продолжается 2-3 мин. Анафаза заканчивается перемещением двух идентичных наборов хромосом (s-хромосом, или одиночных хромосом) к полюсам, где они сближаются, образуя фигуры, напоминающие по внешнему виду (если смотреть со стороны полюса) звезды. Эти фигуры называют дочерними звездами.
Так как хромосомные звезды образуются у каждого из полюсов, данную стадию митоза иногда называют стадией двойной звезды (диастер), или стадией дочерних звезд. Телофаза - конечная стадия митоза, в течение которой на полюсах веретена реконструируются дочерние ядра. Перестройка телофазных хромосом напоминает процессы их изменения в профазе, но происходящие в обратном направлении. При взаимодействии хромосом с мембранными пузырьками цитоплазмы формируется ядерная оболочка. С переходом хромосом в интерфазное состояние образуются новые ядрышки. Телофаза завершается разделением тела клетки - цитотомией, или цитокинезом, что приводит к образованию двух дочерних клеток.
Часть клеток может выходить из цикла репродукции и вступить на путь дифференцировки. Некоторые клетки могут выходить из клеточного цикла в G1-периоде или после S-периода и находиться в покое (Go-период). Такие покоящиеся клетки сохраняют способность к делению и могут снова входить в цикл размножения.
Учебное видео: митоз клетки и его стадии
При проблемах с просмотром скачайте видео со страницы Оглавление темы "Строение клетки. Клеточные элементы.":Временной ход митоза и цитокинеза, типичный для клетки млекопитающего. Точные цифры для разных клеток различны. Цитокинез берёт своё начало в анафазе и завершается, как правило,
к окончанию телофазы
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой. М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую. Первые пять профаза, прометафаза, метафаза, анафаза и телофаза составляют митоз, а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы.
Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны стадии сопряженные с процессами внутриклеточного синтеза: профаза и телофаза. Наиболее быстротечны фазы митоза, в ходе которых происходит движение хромосом: метафаза и анафаза. Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут.
Профаза

К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью.
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом.
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.
Прометафаза

Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
В клетках млекопитающих прометафаза протекает, как правило, в течение 10-20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона около 30 минут.
Метафаза
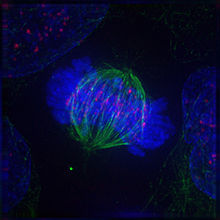
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума.
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.
Анафаза

Анафаза самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью достигающей 0,5-2 мкм/мин., при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10 дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью.
Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно в области кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным.
Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки.
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5-2 раза по сравнению с метафазным. В некоторых других клетках анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным. В растительных клетках анафаза В отсутствует.
Телофаза

Телофаза
Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.
Окончание телофазы преимущественно совпадает с разделением тела материнской клетки цитокинезом. При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы.
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и деление путём образования клеточной пластинки. Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена.
При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократимое кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса.
Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки. После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки.
| Прево, Жан-Луи |